Resti vegetali del Tardo Neogene della “Collezione Coppi” risalente al XIX-XX secolo:
foglie e polline e altri sporomorfi per una ricostruzione paleoambientale.
PRIMA FASE: LO STUDIO DELLE FOGLIE FOSSILI
La prima fase di studio della Collezione Coppi ha comportato l’esame di 78 esemplari di impronte di foglie su limo argilloso che, in alcuni casi, conservano resti di cuticole. L’identificazione è avvenuta mediante osservazioni sulle caratteristiche morfologiche delle foglie: circa il 40% delle impronte fossili è risultata troppo parziale o deteriorata per consentire una identificazione attendibile. I campioni meglio conservati sono stati disegnati con l’aiuto dello stereomicroscopio secondo il metodo della Prof. Zastawniak dello Szafer Institute of Botany di Cracovia, e poi determinati mediante il metodo di Hickey (1973) e il confronto con descrizioni e determinazioni presenti nelle opere degli autori antichi come Massalongo e Scarabelli (1859) e recenti tra i quali Knobloch (1998), Zastawniak (1980), Martinetto (2003) e Kvacek et al. (2004).
Sulla base dell’analisi morfologica e del confronto con le collezioni fossili presenti a Cracovia e al Museo della Terra di Varsavia, le foglie sono state attribuite a sette famiglie: Pinaceae, Taxodiaceae, Corylaceae, Juglandaceae, Rosaceae, Ulmaceae e Fagaceae.
Quest’ultima è risultata la famiglia più rappresentata con 37 esemplari di foglie appartenenti al genere Quercus. In generale, per 29 esemplari le determinazioni di Coppi sono state confermate. Viceversa, in molti casi è stato necessario rivedere le determinazioni proposte da Coppi, ottenendo così nuove attribuzioni tassonomiche. Ad esempio, elementi oggi alloctoni ma presenti nel Neogene, sono risultati Sequoia (Taxodiaceae), Carya e Pterocarya (Juglandaceae), che erano state attribuite da Coppi rispettivamente a Pinaceae e Ulmaceae.
Per giungere a una corretta interpretazione dei risultati ottenuti, è necessario tenere in considerazione che solo una parte degli elementi vegetali che vivevano nell’area è giunta fino a noi come fossile. Ciò dipende da numerosi fattori: la consistenza delle foglie e la loro differente resistenza, l’abbondanza delle diverse specie nel passato, la distanza delle comunità vegetali dalle zone di deposizione, il tipo di trasporto. Per questi ed altri fattori, le associazioni vegetali presenti nel deposito non esauriscono quelle presenti al momento della deposizione: le parti più fragili e le piante più delicate possono mancare, mentre le più coriacee saranno sovrarappresentate (Hummel, 1983). Nel nostro caso, la maggior parte delle filliti appartengono al genere Quercus, latifoglie a foglie caduche che suggerisce la presenza di foreste decidue miste, caratterizzate da esigenze di clima temperato o temperato caldo, o di boschi misti a sempreverdi, ad esempio nelle fasce vegetazionali più elevate, con clima temperato freddo.
Sulla base dell’analisi morfologica e del confronto con le collezioni fossili presenti a Cracovia e al Museo della Terra di Varsavia, le foglie sono state attribuite a sette famiglie: Pinaceae, Taxodiaceae, Corylaceae, Juglandaceae, Rosaceae, Ulmaceae e Fagaceae.
Quest’ultima è risultata la famiglia più rappresentata con 37 esemplari di foglie appartenenti al genere Quercus. In generale, per 29 esemplari le determinazioni di Coppi sono state confermate. Viceversa, in molti casi è stato necessario rivedere le determinazioni proposte da Coppi, ottenendo così nuove attribuzioni tassonomiche. Ad esempio, elementi oggi alloctoni ma presenti nel Neogene, sono risultati Sequoia (Taxodiaceae), Carya e Pterocarya (Juglandaceae), che erano state attribuite da Coppi rispettivamente a Pinaceae e Ulmaceae.
Per giungere a una corretta interpretazione dei risultati ottenuti, è necessario tenere in considerazione che solo una parte degli elementi vegetali che vivevano nell’area è giunta fino a noi come fossile. Ciò dipende da numerosi fattori: la consistenza delle foglie e la loro differente resistenza, l’abbondanza delle diverse specie nel passato, la distanza delle comunità vegetali dalle zone di deposizione, il tipo di trasporto. Per questi ed altri fattori, le associazioni vegetali presenti nel deposito non esauriscono quelle presenti al momento della deposizione: le parti più fragili e le piante più delicate possono mancare, mentre le più coriacee saranno sovrarappresentate (Hummel, 1983). Nel nostro caso, la maggior parte delle filliti appartengono al genere Quercus, latifoglie a foglie caduche che suggerisce la presenza di foreste decidue miste, caratterizzate da esigenze di clima temperato o temperato caldo, o di boschi misti a sempreverdi, ad esempio nelle fasce vegetazionali più elevate, con clima temperato freddo.
SECONDA FASE: LO STUDIO DEL POLLINE FOSSILE

Tre reperti diversi tra loro sono stati scelti per estrarre dall'argilla in cui si sono fossilizzati il polline, e questo per avere ulteriori informazioni sull'ambiente di deposizione e di vita delle piante dell'appennino modenese nel Pliocene.
Negli studi paleoclimatici, il Pliocene è considerato da molti autori un’epoca chiave per la comprensione dell’attuale riscaldamento climatico globale.
La ricerca paleobotanica che, attraverso una rigorosa determinazione dei fossili vegetali porta alla conoscenza della flora passata, offre un contributo in molti casi fondamentale per una ricostruzione attendibile dell’ambiente vegetale e del clima passato. La potenzialità di questa ricerca, però, è funzione del numero dei fossili vegetali scoperti in un determinato territorio. I musei presenti in molte città italiane spesso nascondono un patrimonio da scoprire, collezioni relegate nell’oblio da centinaia di anni in attesa di essere valorizzate e studiate alla luce di nuove conoscenze. In questo contesto, grande interesse assume la collezione di fossili che Francesco Coppi raccolse tra la fine dell’800 e gli inizi del 900 nelle Marne Piacenziane di San Venanzio (Maranello-Modena) nell’Appennino Settentrionale.
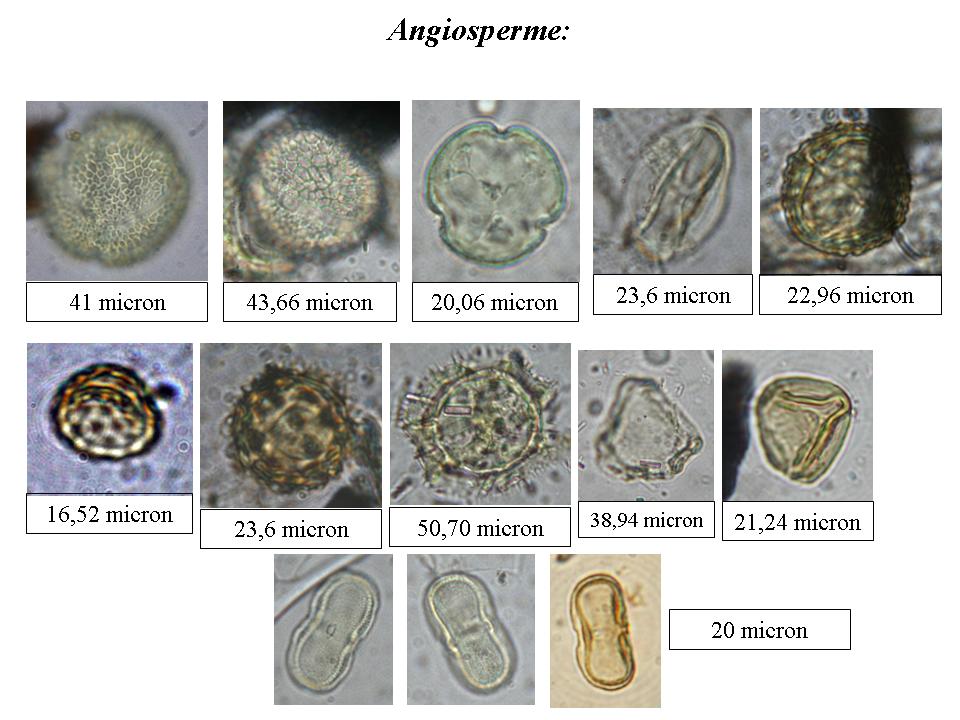
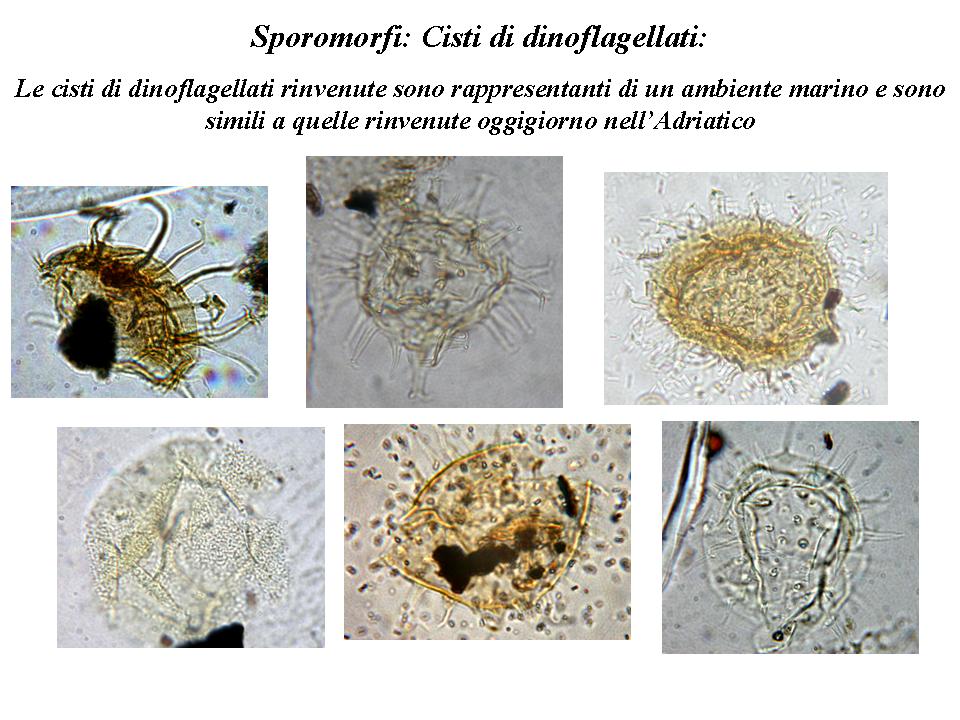
ella seconda fase del lavoro, l’obiettivo è stato quello di isolare i fossili vegetali microscopici, in particolare i granuli pollinici. Con l’aiuto di un piccolo scalpello, prestando attenzione a non recare danno ai fossili, sono stati prelevati frammenti della matrice da tre esemplari di filliti appartenenti a taxa differenti: Algae, Gymnospermae e Angiospermae. I campioni sono stati trattati per concentrare i sporomorfi (cisti algali, spore, polline, ecc…) secondo il metodo di estrazione in uso nel Laboratorio di Palinologia e Paleobotanica dell’Università di Modena e Reggio Emilia. I vetrini fissi allestiti sono stati letti al microscopio ottico a 1000x con immersione in olio. I granuli pollinici sono risultati in generale in ottimo stato di conservazione, e solo in minima quantità deteriorati. Sono stati osservati 1034 granuli pollinici, in prevalenza Gymnospermae (99%). Le Angiospermae appartengono alle famiglie di Compositae, Umbelliferae, Oleaceae, Fagaceae, Gramineae e Rosaceae. La nomenclatura utilizzata è quella messa a punto da Stuchlik (2002). Oltre al polline, sono state osservate un gran numero di cisti di dinoflagellati, che sommati alle alghe presenti tra le filliti, testimoniano un ambiente marino molto simile a quello attuale in Adriatico (Miola in verbis, 2006).
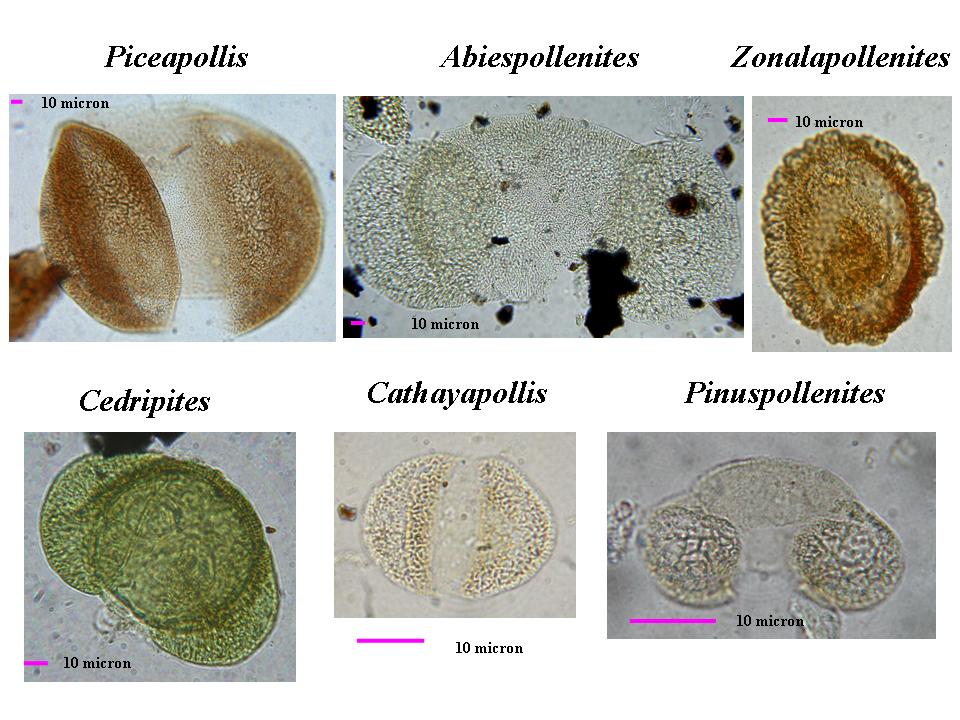
A questo punto è emerso uno degli elementi più nuovi e interessanti della ricerca svolta sulla Collezione Coppi: il polline appartiene per lo più a taxa diversi da quelli rappresentati dalle filliti. Gli sporomorfi, inclusi nel sedimento in cui si sono fossilizzate le nostre filliti, portano, infatti, alla ricostruzione di un ambiente dominato da conifere come pini e abeti (Abies, Picea e Tsuga), molto differente da quello mostrato dai macrofossili dominato, come detto sopra, da caducifoglie. Per chiunque si occupi di studio del polline presente in carote marine questo dato non è una sorpresa: è noto, infatti, che nei depositi marini il polline di conifere (e in particolare quello provvisto di “sacche”, una morfologia che ne facilita il galleggiamento in acqua) è sovrarappresentato e indica provenienza anche da centinaia di chilometri. Proprio questa apparente contraddizione suggerisce interessanti informazioni per la ricostruzione paleoambientale:
-le associazioni vegetali indicate dai microfossili rappresentano in prevalenza le foreste di conifere che dovevano essere distribuite nelle fasce altitudinali più elevate, anche molto lontane dal sito di deposizione;
-i macrofossili, invece, rappresentano la vegetazione a latifoglie decidue, con diffusi boschi di querce, più prossima al luogo della loro fossilizzazione.
Per capire il significato di questi dati pollinici non è forse superfluo ricordare che, in questa ricerca, non è stata condotta la classica analisi pollinica di una successione diacronica dove campioni in successione descrivono oscillazioni vegetazionali/climatiche. È stata invece eseguita un’analisi di campioni isolati, forse il primo tentativo del genere che sicuramente sarà da testare con altri casi, ma che ha già prodotto interessanti primi risultati.
Dai campioni provenienti dalla matrice di tre macrofossili diversi Algae, Gymnospermae e Angiospermae, infatti, sono emersi dati micropaleobotanici assai simili tra loro (cioè, gli assemblaggi pollinici dei tre campioni sono somiglianti): ciò consente di confermare che i macrofossili studiati devono essersi depositati nello stesso luogo e nello stesso tempo. Dalla integrazione dei taxa pollinici con quelli delle foglie, inoltre, emerge la complessità di un paesaggio che deve essere immaginato, allora come ora, composito e distribuito su più piani vegetazionali.
Riassumendo, l’insieme delle informazioni estrapolabili da questi dati paleobotanici, pur tenendo presente la casualità dell’evento di fossilizzazione che potrebbe aver portato a una conservazione selettiva delle foglie o dei pollini più resistenti, sono preziose per dare un’istantanea dell’ambiente presente al momento della deposizione dei fossili raccolti dal Coppi, collocabili in un momento del Neogene sull’Appennino Emiliano. Tale ambiente, anche in accordo con i dati geologici (Gasperi et al., 1989; Gasperi, 1995), era per lo più di confine tra ambiente marino e continentale.
Negli studi paleoclimatici, il Pliocene è considerato da molti autori un’epoca chiave per la comprensione dell’attuale riscaldamento climatico globale.
La ricerca paleobotanica che, attraverso una rigorosa determinazione dei fossili vegetali porta alla conoscenza della flora passata, offre un contributo in molti casi fondamentale per una ricostruzione attendibile dell’ambiente vegetale e del clima passato. La potenzialità di questa ricerca, però, è funzione del numero dei fossili vegetali scoperti in un determinato territorio. I musei presenti in molte città italiane spesso nascondono un patrimonio da scoprire, collezioni relegate nell’oblio da centinaia di anni in attesa di essere valorizzate e studiate alla luce di nuove conoscenze. In questo contesto, grande interesse assume la collezione di fossili che Francesco Coppi raccolse tra la fine dell’800 e gli inizi del 900 nelle Marne Piacenziane di San Venanzio (Maranello-Modena) nell’Appennino Settentrionale.
ella seconda fase del lavoro, l’obiettivo è stato quello di isolare i fossili vegetali microscopici, in particolare i granuli pollinici. Con l’aiuto di un piccolo scalpello, prestando attenzione a non recare danno ai fossili, sono stati prelevati frammenti della matrice da tre esemplari di filliti appartenenti a taxa differenti: Algae, Gymnospermae e Angiospermae. I campioni sono stati trattati per concentrare i sporomorfi (cisti algali, spore, polline, ecc…) secondo il metodo di estrazione in uso nel Laboratorio di Palinologia e Paleobotanica dell’Università di Modena e Reggio Emilia. I vetrini fissi allestiti sono stati letti al microscopio ottico a 1000x con immersione in olio. I granuli pollinici sono risultati in generale in ottimo stato di conservazione, e solo in minima quantità deteriorati. Sono stati osservati 1034 granuli pollinici, in prevalenza Gymnospermae (99%). Le Angiospermae appartengono alle famiglie di Compositae, Umbelliferae, Oleaceae, Fagaceae, Gramineae e Rosaceae. La nomenclatura utilizzata è quella messa a punto da Stuchlik (2002). Oltre al polline, sono state osservate un gran numero di cisti di dinoflagellati, che sommati alle alghe presenti tra le filliti, testimoniano un ambiente marino molto simile a quello attuale in Adriatico (Miola in verbis, 2006).
A questo punto è emerso uno degli elementi più nuovi e interessanti della ricerca svolta sulla Collezione Coppi: il polline appartiene per lo più a taxa diversi da quelli rappresentati dalle filliti. Gli sporomorfi, inclusi nel sedimento in cui si sono fossilizzate le nostre filliti, portano, infatti, alla ricostruzione di un ambiente dominato da conifere come pini e abeti (Abies, Picea e Tsuga), molto differente da quello mostrato dai macrofossili dominato, come detto sopra, da caducifoglie. Per chiunque si occupi di studio del polline presente in carote marine questo dato non è una sorpresa: è noto, infatti, che nei depositi marini il polline di conifere (e in particolare quello provvisto di “sacche”, una morfologia che ne facilita il galleggiamento in acqua) è sovrarappresentato e indica provenienza anche da centinaia di chilometri. Proprio questa apparente contraddizione suggerisce interessanti informazioni per la ricostruzione paleoambientale:
-le associazioni vegetali indicate dai microfossili rappresentano in prevalenza le foreste di conifere che dovevano essere distribuite nelle fasce altitudinali più elevate, anche molto lontane dal sito di deposizione;
-i macrofossili, invece, rappresentano la vegetazione a latifoglie decidue, con diffusi boschi di querce, più prossima al luogo della loro fossilizzazione.
Per capire il significato di questi dati pollinici non è forse superfluo ricordare che, in questa ricerca, non è stata condotta la classica analisi pollinica di una successione diacronica dove campioni in successione descrivono oscillazioni vegetazionali/climatiche. È stata invece eseguita un’analisi di campioni isolati, forse il primo tentativo del genere che sicuramente sarà da testare con altri casi, ma che ha già prodotto interessanti primi risultati.
Dai campioni provenienti dalla matrice di tre macrofossili diversi Algae, Gymnospermae e Angiospermae, infatti, sono emersi dati micropaleobotanici assai simili tra loro (cioè, gli assemblaggi pollinici dei tre campioni sono somiglianti): ciò consente di confermare che i macrofossili studiati devono essersi depositati nello stesso luogo e nello stesso tempo. Dalla integrazione dei taxa pollinici con quelli delle foglie, inoltre, emerge la complessità di un paesaggio che deve essere immaginato, allora come ora, composito e distribuito su più piani vegetazionali.
Riassumendo, l’insieme delle informazioni estrapolabili da questi dati paleobotanici, pur tenendo presente la casualità dell’evento di fossilizzazione che potrebbe aver portato a una conservazione selettiva delle foglie o dei pollini più resistenti, sono preziose per dare un’istantanea dell’ambiente presente al momento della deposizione dei fossili raccolti dal Coppi, collocabili in un momento del Neogene sull’Appennino Emiliano. Tale ambiente, anche in accordo con i dati geologici (Gasperi et al., 1989; Gasperi, 1995), era per lo più di confine tra ambiente marino e continentale.
Bibliografia
- Coppi F. (1869). Catalogo dei fossili miocenici e pliocenici del Modenese. Annuario della Società dei Naturalisti di Modena, IV, 163-228, Modena
- Gasperi G., Cremaschi M., Mantovani Uguzzoni M.P., Cardarelli A., Cattani M., Labate D. (1987). Evoluzione Plio-Quaternaria del margine Appenninico Modenese e dell’antistante pianura. Note illustrative alla carta geologica – Estratto da Mem. Soc. Geol.It, 375 – 431, 8ff., stab., 1 carta geologica (Società Geologica Italiana, 1989)
- Hickey L. J. (1973). Classification of the architecture of Dicotyledonous Leaves - American Journal of Botany Vol. 60 No.1 83 pp. January 1973
- Hummel A. (1983). The Pliocene leaf flora from Ruszòw near Zary in Lower Silesia, SW Poland. Prace Museum Ziemi. Zeszyt. 36 Warszawa 1983. 9-99, Prace Paleobotaniczne
- Knobloch E. (1998). Der Pliozane Laubwald Von Willershausen Am Harz (Mitteleuropa). 102 pp. Documenta naturae n° 120, Munchen
- Kwaček Z. Dvořák Z., Mach K., Sakala J.(2004). Třetihorní rostlini. 158 pp. Severočeské doly a.s., Chomutov Granit, s.r.o. Praha
- Martinetto E. (2003). Leaves of terrestrial Plants from the Pliocene shallow marine and transitional deposits of Asti (Piedmont, Italy). Bollettino della Società Paleontologica Italiana, 42 (1-2),2003 75-111
- Massalogno A., Scarabelli E. (1859). Studii sulla Flora Fossile e Geologia stratigrafica del Senigalliese. 167 pp. Tipografia Galeati e Figlio, Imola
- Stuchlik L. (2002). Atlas of pollen and spores of the Polish Neogene - Volume 2 Gymnosperms 237 pp. Edited by L. Stuchlik
- Zastawniak E. (1980). Sarmatian leaf flora from the southern margin of the Holy Cross MTS. South Poland. Prace Museum Ziemi Zeszyt 33, 33 - 105 Warsawa. Prace Paleobotaniczne
PER SAPERNE DI PIU'.....
Palinologia e microscopia elettronica
scritto dalla Prof. Brugiapaglia Elisabetta – Dipartimento SAVA Facoltà di Agraria
Le specie vegetali appartenenti alle Tracheophyta producono quantità spesso rilevanti di polline e spore che, viventi o fossili, sono l’oggetto di studio principale della Palinologia. Questa scienza si occupa in senso lato anche di altri sporomorfi quali, ad esempio, cisti di dinoflagellati, altri elementi algali, elementi fungini e spore di Briofite.
Il termine “Palinologia” fu proposto per la prima volta nel 1944 dai botanici inglesi Hyde e Williams per indicare la ‘scienza delle particelle polverulente - polline e altri palinomorfi - galleggianti in aria’. La parola è ispirata al verbo greco palunein - palynein = diffondere, disperdere, che ha assonanza con il greco paipále e il latino pollen, lemmi traducibili entrambi in ‘fior di farina’.
Gli antichi conoscevano empiricamente il ruolo del polline di alcune piante utili e attuavano l’impollinazione artificiale. Ne sono testimonianza i testi cuneiformi di Ur (2300 a.C.; Wrigley, 1995) e i bassorilievi del palazzo del re assiro Ashuirnasirpal II (895-883 a.C.) che rappresentano figure mistiche nell’atto di agitare le infiorescenze maschili della palma da dattero sopra quelle femminili (Pacini, 1987).
Bisogna però arrivare al secolo XVI perché l’idea che le piante abbiamo due sessi caratterizzati da strutture distinte cominci ad essere accettata. Alla fine del XVII secolo, Camerarius dimostra che l’ovulo di una pianta non può svilupparsi in seme senza essere stato prima sottoposto all’azione del polline che si trova sugli stami, sesso maschile della pianta (Pons, 1970).
Solo nella seconda metà del XVII secolo, grazie alla scoperta del microscopio ottico, diventa possibile intraprendere i primi studi sulla morfologia dei granuli pollinici. Le prime descrizioni corredate da disegni sono fornite quasi contemporaneamente dall’inglese Grew e dall’italiano Malpighi (Accorsi et al., 2005). Nel XVIII secolo, Kolreuter, Sprengel e Amici svolgono studi sulla fecondazione. All’inizio del XIX secolo, Bauer, pittore botanico alla corte del re Giorgio III, realizza il disegno del polline di 175 specie riconoscendo le loro forme essenziali. Purkinje, nel 1830, caratterizza il polline delle principali famiglie e, grazie al progresso tecnico del microscopio, nel 1834 Von Mohl realizza una classificazione dei tipi pollinici. Alla fine del XIX secolo, Fischer pone le basi per le attuali conoscenze della parete pollinica.
La nascita dell’analisi pollinica, cioè l’analisi statistica del contenuto di polline e spore in un substrato, è generalmente attribuita agli studi di von Post, geologo svedese che nel 1916 presenta i primi diagrammi pollinici ottenuti da sedimenti (Faegri et al., 1989). Nel 1934, Raistrick dimostra la possibilità di utilizzare la palinologia a fini stratigrafici, consolidando uno dei settori più prolifici della palinologia applicata allo studio di substrati del passato o Paleopalinologia. Negli stessi anni, Firbas intuisce la possibilità di rintracciare nei diagrammi pollinici influenze antropiche sulla vegetazione, un aspetto che porterà in tempi più recenti allo sviluppo della Archeopalinologia o studio del polline in siti antropizzati.
Nel 1835 Kirkman, essendo lui stesso allergico, era riuscito ad individuare la natura del polline responsabile del raffreddore da fieno, un disturbo genericamente conosciuto sin dal XVI secolo. Da quelle prime osservazioni si sarebbe poi sviluppata nel XX secolo la Aeropalinologia o studio del polline diffuso in aria.
Nel 1895 Pfister aveva scritto il primo lavoro sull'analisi pollinica dei mieli, mettendo in relazione lo spettro pollinico del miele con il suo odore, il suo gusto e la sua origine botanica. In quegli anni vari studiosi iniziarono a dedicarsi a questo particolare ramo della palinologia, la Melissopalinologia, che ha avuto un ruolo fondamentale per la caratterizzazione, commercializzazione e controllo di qualità dei mieli, nonché per la conoscenza della flora apistica e del comportamento delle api.
La Palinologia può essere suddivisa in vari settori: a) Aeropalinologia, polline e spore attuali diffusi in aria, b) Briopalinologia, polline attuale catturato nei muschi, licheni e altre piante a cuscinetto che fungono da trappole naturali, c) Melissopalinologia, polline nei mieli e in altri prodotti apistici, d) Farmacopalinologia, polline nei prodotti erboristici e droghe in genere, e) Bromatopalinologia, polline negli alimenti, f) Copropalinologia, polline negli escrementi; g) Criminopalinologia, o Palinologia Forense, polline in substrati vari legati a indagini forensi, h) Speleopalinologia, polline e spore conservati in grotta; i) Paleopalinologia, polline e spore fossili conservati in depositi marini, lacustri, torbiere, suoli, l) Archeopalinologia, in siti interessati da presenza antropica.
Il polline presenta una parete (esina) estremamente complessa con una struttura e delle sculture composte da elementi singoli quali columelle, gemmule, spine e verruche. Questi elementi rendono il polline estremamente specifico e identificabile facolmente soprattutto a livello di genere.
La migliore metodologia di osservazione per l’analisi della struttura (comprendente l’insieme delle caratteristiche interne delle columelle e del tetto) e delle sculture (comprendente le ornamentazioni geometriche esterne all’esina), è la cosidetta “illuminazione di Köhler”: si effettua con il microscopio ottico a luce trasmessa osservando il granulo secondo il metodo della L.O. analisi (Luce-Oscuro) che consiste nell’osservare nel centro del campo del microscopio una piccola superficie dell’esina, assimilabile ad un piano, nella quale si realizzano diverse messe a fuoco in tutto lo spessore dell’esina, dalla sommità delle sculture fino all’endesina; con questo metodo, un elemento in rilievo apparirà prima luminoso poi oscuro (Bertrand, 1961; Reille, 1990).
Un altro modo di osservare i granuli pollinici è attraverso la microscopia elettronica, grazie alla quale è stato possibile studiare la superficie dell’esina
Il riconoscimento e le osservazioni di routine (conteggi) vengono effettuati con il microscopio ottico regolato secondo l’analisi L-O (luce-ombra).
Il microscopio elettronico offre delle performance altamente specifiche soprattutto per richerche di carattere morfologico e per risolvere problemi tassonomici in gruppi vegetali particolarmente complessi.
Le due tipologie di osservazione sono assolutamente complementari in quanto hanno un campo d’indagine dello stesso oggetto differenziato: in particolare con la microscopia ottica possiamo analizzare lo spessore, le aperture dell’esina mentre con il microscopio elettronico osserviamo le sculture presenti sull’esina. L’estrema precisione del microscopio elettronico è quindi fondamentale per le ricerche tassonomiche ma rappresenterebbe un impedimento nell’analisi di conteggio di routine.
Le osservazioni realizzate presso il centro di microscopia elettronica dell’Università del Molise hanno avuto come scopo principale quello di rappresentare le diverse tipologie di esina nell’ambito di una ricerca che verrà a breve pubblicata in un testo di botanica per gli studenti universitari.
scritto dalla Prof. Brugiapaglia Elisabetta – Dipartimento SAVA Facoltà di Agraria
Le specie vegetali appartenenti alle Tracheophyta producono quantità spesso rilevanti di polline e spore che, viventi o fossili, sono l’oggetto di studio principale della Palinologia. Questa scienza si occupa in senso lato anche di altri sporomorfi quali, ad esempio, cisti di dinoflagellati, altri elementi algali, elementi fungini e spore di Briofite.
Il termine “Palinologia” fu proposto per la prima volta nel 1944 dai botanici inglesi Hyde e Williams per indicare la ‘scienza delle particelle polverulente - polline e altri palinomorfi - galleggianti in aria’. La parola è ispirata al verbo greco palunein - palynein = diffondere, disperdere, che ha assonanza con il greco paipále e il latino pollen, lemmi traducibili entrambi in ‘fior di farina’.
Gli antichi conoscevano empiricamente il ruolo del polline di alcune piante utili e attuavano l’impollinazione artificiale. Ne sono testimonianza i testi cuneiformi di Ur (2300 a.C.; Wrigley, 1995) e i bassorilievi del palazzo del re assiro Ashuirnasirpal II (895-883 a.C.) che rappresentano figure mistiche nell’atto di agitare le infiorescenze maschili della palma da dattero sopra quelle femminili (Pacini, 1987).
Bisogna però arrivare al secolo XVI perché l’idea che le piante abbiamo due sessi caratterizzati da strutture distinte cominci ad essere accettata. Alla fine del XVII secolo, Camerarius dimostra che l’ovulo di una pianta non può svilupparsi in seme senza essere stato prima sottoposto all’azione del polline che si trova sugli stami, sesso maschile della pianta (Pons, 1970).
Solo nella seconda metà del XVII secolo, grazie alla scoperta del microscopio ottico, diventa possibile intraprendere i primi studi sulla morfologia dei granuli pollinici. Le prime descrizioni corredate da disegni sono fornite quasi contemporaneamente dall’inglese Grew e dall’italiano Malpighi (Accorsi et al., 2005). Nel XVIII secolo, Kolreuter, Sprengel e Amici svolgono studi sulla fecondazione. All’inizio del XIX secolo, Bauer, pittore botanico alla corte del re Giorgio III, realizza il disegno del polline di 175 specie riconoscendo le loro forme essenziali. Purkinje, nel 1830, caratterizza il polline delle principali famiglie e, grazie al progresso tecnico del microscopio, nel 1834 Von Mohl realizza una classificazione dei tipi pollinici. Alla fine del XIX secolo, Fischer pone le basi per le attuali conoscenze della parete pollinica.
La nascita dell’analisi pollinica, cioè l’analisi statistica del contenuto di polline e spore in un substrato, è generalmente attribuita agli studi di von Post, geologo svedese che nel 1916 presenta i primi diagrammi pollinici ottenuti da sedimenti (Faegri et al., 1989). Nel 1934, Raistrick dimostra la possibilità di utilizzare la palinologia a fini stratigrafici, consolidando uno dei settori più prolifici della palinologia applicata allo studio di substrati del passato o Paleopalinologia. Negli stessi anni, Firbas intuisce la possibilità di rintracciare nei diagrammi pollinici influenze antropiche sulla vegetazione, un aspetto che porterà in tempi più recenti allo sviluppo della Archeopalinologia o studio del polline in siti antropizzati.
Nel 1835 Kirkman, essendo lui stesso allergico, era riuscito ad individuare la natura del polline responsabile del raffreddore da fieno, un disturbo genericamente conosciuto sin dal XVI secolo. Da quelle prime osservazioni si sarebbe poi sviluppata nel XX secolo la Aeropalinologia o studio del polline diffuso in aria.
Nel 1895 Pfister aveva scritto il primo lavoro sull'analisi pollinica dei mieli, mettendo in relazione lo spettro pollinico del miele con il suo odore, il suo gusto e la sua origine botanica. In quegli anni vari studiosi iniziarono a dedicarsi a questo particolare ramo della palinologia, la Melissopalinologia, che ha avuto un ruolo fondamentale per la caratterizzazione, commercializzazione e controllo di qualità dei mieli, nonché per la conoscenza della flora apistica e del comportamento delle api.
La Palinologia può essere suddivisa in vari settori: a) Aeropalinologia, polline e spore attuali diffusi in aria, b) Briopalinologia, polline attuale catturato nei muschi, licheni e altre piante a cuscinetto che fungono da trappole naturali, c) Melissopalinologia, polline nei mieli e in altri prodotti apistici, d) Farmacopalinologia, polline nei prodotti erboristici e droghe in genere, e) Bromatopalinologia, polline negli alimenti, f) Copropalinologia, polline negli escrementi; g) Criminopalinologia, o Palinologia Forense, polline in substrati vari legati a indagini forensi, h) Speleopalinologia, polline e spore conservati in grotta; i) Paleopalinologia, polline e spore fossili conservati in depositi marini, lacustri, torbiere, suoli, l) Archeopalinologia, in siti interessati da presenza antropica.
Il polline presenta una parete (esina) estremamente complessa con una struttura e delle sculture composte da elementi singoli quali columelle, gemmule, spine e verruche. Questi elementi rendono il polline estremamente specifico e identificabile facolmente soprattutto a livello di genere.
La migliore metodologia di osservazione per l’analisi della struttura (comprendente l’insieme delle caratteristiche interne delle columelle e del tetto) e delle sculture (comprendente le ornamentazioni geometriche esterne all’esina), è la cosidetta “illuminazione di Köhler”: si effettua con il microscopio ottico a luce trasmessa osservando il granulo secondo il metodo della L.O. analisi (Luce-Oscuro) che consiste nell’osservare nel centro del campo del microscopio una piccola superficie dell’esina, assimilabile ad un piano, nella quale si realizzano diverse messe a fuoco in tutto lo spessore dell’esina, dalla sommità delle sculture fino all’endesina; con questo metodo, un elemento in rilievo apparirà prima luminoso poi oscuro (Bertrand, 1961; Reille, 1990).
Un altro modo di osservare i granuli pollinici è attraverso la microscopia elettronica, grazie alla quale è stato possibile studiare la superficie dell’esina
Il riconoscimento e le osservazioni di routine (conteggi) vengono effettuati con il microscopio ottico regolato secondo l’analisi L-O (luce-ombra).
Il microscopio elettronico offre delle performance altamente specifiche soprattutto per richerche di carattere morfologico e per risolvere problemi tassonomici in gruppi vegetali particolarmente complessi.
Le due tipologie di osservazione sono assolutamente complementari in quanto hanno un campo d’indagine dello stesso oggetto differenziato: in particolare con la microscopia ottica possiamo analizzare lo spessore, le aperture dell’esina mentre con il microscopio elettronico osserviamo le sculture presenti sull’esina. L’estrema precisione del microscopio elettronico è quindi fondamentale per le ricerche tassonomiche ma rappresenterebbe un impedimento nell’analisi di conteggio di routine.
Le osservazioni realizzate presso il centro di microscopia elettronica dell’Università del Molise hanno avuto come scopo principale quello di rappresentare le diverse tipologie di esina nell’ambito di una ricerca che verrà a breve pubblicata in un testo di botanica per gli studenti universitari.

Isypedia by Isabella Massamba is licensed under a Creative Commons Attribution-NonCommercial-NoDerivs 3.0 Unported License.
